
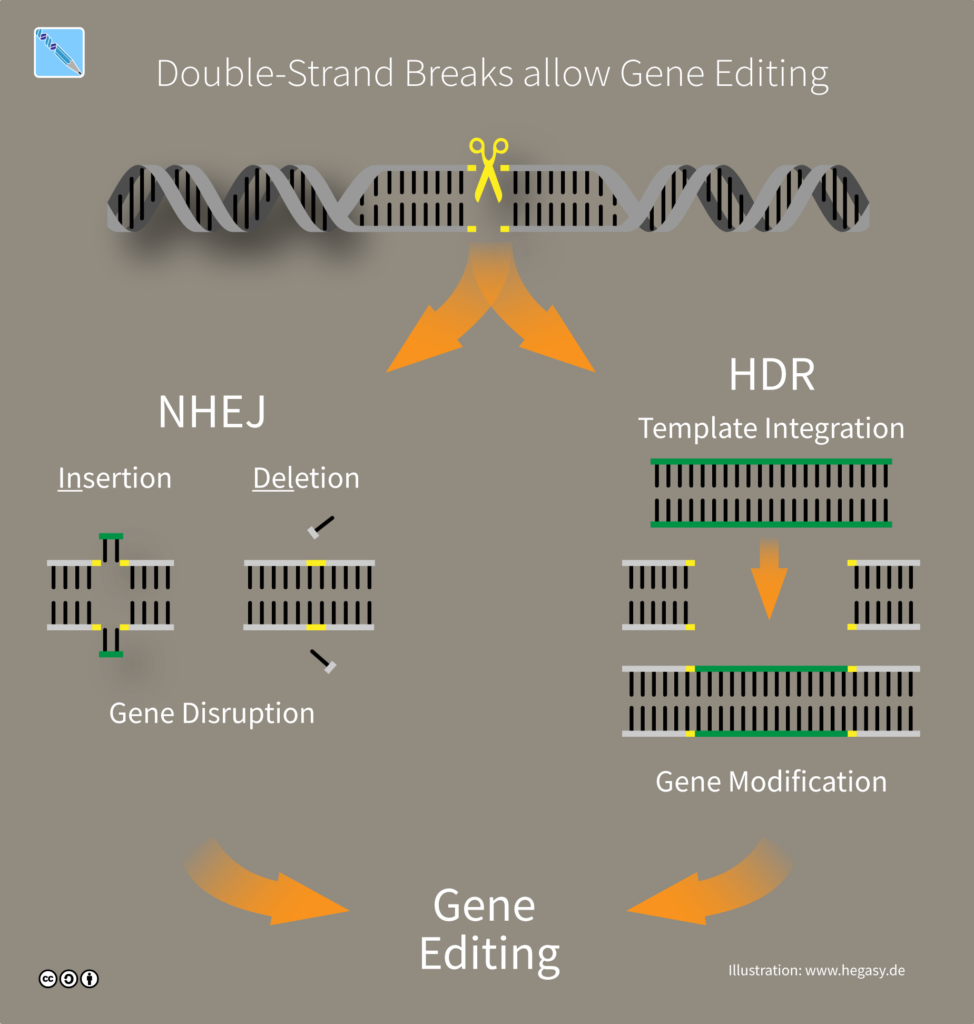
Las células cuentan con distintos mecanismos para reparar rupturas de doble cadena en el ADN (double-strand breaks, DSBs). Entre los principales se encuentran dos vías: Non-Homologous End Joining (NHEJ) y Homology-Directed Repair (HDR) [1, 2]. Ambos procesos son esenciales para la supervivencia celular, pero difieren en su precisión, sus requerimientos y su relevancia en edición génica.
Non-Homologous End Joining (NHEJ) – Unión de extremos no homólogos
Es el mecanismo de reparación más frecuente, en el cual ciertas proteínas forman complejos que se unen los extremos del ADN para protegerlo y evitar su degradación, hasta que una ligasa de ADN cierra la ruptura catalizando la formación de enlaces covalentes entre esos extremos.
Este es un proceso muy rápido que, al no utilizar ninguna plantilla de reparación, puede llegar a inducir errores como inserciones o deleciones (indels) en el sitio de la ruptura, pudiendo alterar la secuencia original del ADN. Por esta razón, se dice que es un mecanismo “error-prone” o propenso a errores. La edición génica capitaliza este “error” para promover la aparición de indels en un lugar específico. Por ejemplo, si se produce un indel en una secuencia codificante, esto puede conducir a la disrupción del marco de lectura abierto (open reading frame, ORF) y generar un codón stop prematuro que impida su transcripción. Esto se conoce como knock-out génico y permite inducir la pérdida de función de un gen.
Si hablamos de “error” es porque la generación de indels es infrecuente y la mayoría de las reparaciones por NHEJ van a mantener la secuencia original. Sin embargo, si el complejo gRNA-Cas9 se mantiene constitutivamente expresado y dirigido al sitio blanco, los ciclos de DSB y reparación continuarán ocurriendo, aumentando la probabilidad de cambios en el ADN. Mientras que el sitio “editado” ya no será susceptible de edición, pues perderá la complementariedad con el gRNA, los productos de reparación que mantuvieron la secuencia original volverán a ser blanco de Cas9, favoreciendo así la acumulación de alelos editados.
Homology-Directed Repair (HDR) – Reparación dirigida por homología
Este mecanismo es menos frecuente pero altamente preciso, porque utiliza una molécula de ADN homólogo como plantilla de reparación (repair template, RT) para corregir el daño. Por eso, en lugar de ser “error-prone”, se considera “error-free”.
El proceso comienza con la resección de los extremos rotos por parte de exonucleasas, lo que genera regiones de cadena simple que pueden hibridarse con una plantilla homóloga, típicamente la cromátida hermana. De esta manera, se forma una estructura de ADN de cadena doble, donde una de las cadenas rotas es “completada” por una polimerasa de ADN con la información proporcionada. Después de que la cadena ha sido reparada, la estructura de ADN se resuelve y es ligado por enzimas ligasas, quedando completamente restaurado.
La reparación por HDR es mucho menos frecuente porque depende de la fase del ciclo celular. En condiciones “normales”, es decir, sin intervención de edición génica, el mecanismo HDR es más activo en las fases S (síntesis de ADN, donde se replica el genoma) y G2 (fase de preparación para la división celular), cuando la cromátida hermana está disponible como plantilla de reparación. En cambio, en la fase G1 (crecimiento celular previo a la replicación), esta plantilla no está presente, por lo que predomina mayoritariamente NHEJ, que es más rápido y no requiere homología. Durante la fase M (mitosis), la reparación del ADN se encuentra fuertemente restringida debido al alto grado de condensación cromosómica, por lo que también prevalecen mecanismos rápidos como NHEJ.
En edición genética, el mecanismo HDR puede ser utilizado para realizar cambios muy precisos, como knockins y reemplazos alélicos, pues la plantilla de reparación se puede diseñar “a medida” para que contenga e incorpore los cambios deseados. A diferencia de un “knock-out“, que implica la eliminación o desactivación de un gen, un “knock-in” implica la introducción de un nuevo material genético, como un gen de interés, una secuencia reguladora, o una mutación específica, en un sitio específico del genoma. Sin embargo, la tasa de edición alcanzada será mucho más baja que con NHEJ, porque la plantilla de reparación debe coincidir en tiempo y espacio con el sitio de la ruptura del ADN e incluso así, la mayoría de las reparaciones ocurrirán mediante NHEJ, que es el patrón de reparación dominante.
Por esta razón, las publicaciones científicas actuales reportan la edición génica de plantas mayoritariamente a través del mecanismo NHEJ, donde las herramientas CRISPR se utilizan principalmente como un mutágeno biológico que inducen el corrimiento del marco en genes de interés. Sin embargo, se han desarrollado estrategias alternativas como el base editing y el prime editing, las cuales permiten realizar cambios precisos sin depender de rupturas de doble cadena ni de recombinación homóloga.
https://commons.wikimedia.org/wiki/File:16_Hegasy_DNA_Rep_Wiki_E_CCBYSA.png
{kind=link}